Complementary Correlational Profiles between Slow Wave Brain Oscillations and Working Memory Capacity
A B S T R A C T
Working memory involves a range of functions, including maintenance of information and processing that information undisturbed by distraction. Neuroscientific studies have observed critical contributions from frontal and parietal brain regions during processing of cognitive demanding tasks. However, less is known about individual differences in the resting state and their association with working memory capacity. In this study, electrophysiological recordings were taken from thirty volunteers in eyes closed and eyes open conditions after completing the automated version of the operation span task. The results reveal two clusters of correlations: a midline-theta cluster and a parieto-temporal alpha cluster. The theta and alpha clusters have a negative and a positive correlation with operation span performance, respectively. These results are interpreted as individual differences in cognitive preparedness, with the centro-parietal region being critical in switching between outward and inward attention, with the balance of theta and alpha spectral power at Pz indicating to where cognitive resources are directed.
Keywords
Brain oscillations, alpha rhythm, theta rhythm, working memory capacity, operation span task
Introduction
Working memory is one of the most investigated topics in cognitive and neural sciences. Its definition has been adjusted in relation to new findings and insights and over fifteen different theories exist about working memory and its characteristics [1]. Neuroscientists have contributed to this field both at the level of theory building and empirically by searching for the neural substrate of working memory. Of particular interest is the capacity of working memory. In some theories, the capacity is time-limited whereas other theories assume an item-based limitation [2-4]. Yet others either assume that both time and item-based limitations apply or that no limitation exist at all [5-7]. Because of the lack of consensus about capacity limitations, the interpretations of performance on so-called memory span tasks also lack consensus.
In order to pursue a common understanding, the field has focused on processes that are carried out within working memory. This approach has led to findings of neural substrates involved in working memory processing. For example, parietal regions are involved in phonological processing in working memory tasks [8-10]. Shivde and Thompson-Schill used a delayed judgement task to probe the neural substrates of different levels of processing during the delay interval [11]. They observed that phonological processing is supported by the left superior parietal gyrus and demonstrated that semantic maintenance was supported by bilateral inferior frontal gyrus and the left middle temporal gyrus. In the visual domain, in a visual working memory task, a slow negative wave increased in amplitude when the memory load increases [12, 13]. In a meta-analysis, it was shown that there is a significant and sizeable correlation between the EEG pattern and working memory capacity [14]. At present there are no studies that takes performance of a span task and ask with what EEG profile it correlates. The present study aims to do this using the operation span task and EEG.
The operation span task is a prototypical working memory task that requires processing of operations and maintenance of information [15]. Neuroscientific studies that used the operation span task have grouped people into high or low span and investigate the neural differences on other tasks. For example, using fMRI, Osaka and colleagues observed increased activation in inferior frontal, mid-frontal and parietal regions during performance of the operation span task [16]. Scharinger and colleagues found a decrease in alpha and an increase in theta brain wave at fronto-central and parieto-occipital during performance of the operation span task [17]. This was also observed in other working memory tasks [18, 19]. Increasing demands on working memory have been found to increase alpha band power, but most of these studies measure the EEG when the participants are engaged in processing a task, varying the working memory demand [20]. This opens up the possibility that any idiosyncratic strategies used during the task could contaminate the EEG recordings, resulting in ambiguous results across studies.
Scalp-recorded theta oscillations may be due to local cortical generators, but can also be driven by deeper sources, such as the hippocampi that are critical for memory processing. These rhythms are ideally suited for integrating local and global information, a function of working memory. The aim of the current study is to correlate performance on the operation span task with EEG brain wave oscillations measured at rest. The assumption is that resting-state measurements are uncontaminated with the various processing that occurs during task performance and therefore provide a stable reference point for measuring correlations. The data for this study comes from a recent article in which 30 participants’ brain waves were recorded for 5 minutes in rest with eyes open and eyes closed [21]. The methods of the study are in the original article, but a summary of the main relevant features is presented next.
Materials and Methods
A total of 30 participants (mean age = 31 years, SD = 9.4) were tested on the operation span task and EEG recordings. The operation span task required participants to memorise letters and solving mathematical operations [22]. In the task, participants saw an operation such as “6/2 - 3 =” and indicate the answer. Immediately after this, they would see a letter that they had to commit to memory. A second operation-letter pair was then presented. This repeated until a given number of operation-letter pairs were presented (the span length), which is between 3 and 7. After the final operation-letter pair, the participant was asked to report all letters in the order in which they were shown. The score on the operation span task was the total number of correctly remembered letters (maximum sore is 75). The EEG recordings used 19 channels, referenced to the auricles, at a sampling rate of 256 Hz. Impedance was below 10k Ohm for all electrodes. Participants were seated in a reclining armchair with leg support in a 45 degrees angle. Their brain waves were recorded for five minutes with eyes open and then for five minutes with eyes closed.
Results
Working memory capacity varied among participants. The mean was 42.8 (SD = 13.4) and all participants obtained a score between the minimum and maximum possible score. The OSPAN scores were correlated with the relative power spectrum in all frequency bins across all electrodes. The unthresholded correlations are shown in the top row of (Figure 1), with eyes closed (left panel) and eyes open (right panel). The black lines demarcate the five regions across the anterior-posterior axis. A clear pattern emerges in which working memory capacity has a negative correlation with spectral power at 3-4 Hz (low theta) and a positive correlation with spectral power at 8-10 Hz (low alpha). This pattern is similar across all electrodes.
Given the exploratory nature of this analysis, a permutation test was conducted. More specifically, the individual capacities were randomly shuffled 10,000 times and the resulting random correlations with the EEG data were recorded. This produced a null distribution of 7,600,000 correlations, which was used to obtain the permutation p-values of each of the unthresholded correlation. A two-tailed test was applied with a significance level of 0.05 (0.025 at either side of the null-distribution). The second row in (Figure 1) shows the thresholded correlations in both eyes closed and open conditions, i.e., the correlations that remained significant under the global permutation null distribution. This reveals that only a subset of electrodes and frequencies remain significantly correlated with operation span scores.
In a final step, only those electrode-frequency pairs that remained significant under both eyes closed and eyes open conditions were retained and shown in the bottom row of (Figure 1) as topographic maps, excluding 29-31 Hz at O2, where the correlations were less than 0.4 for both ocular conditions. The results show that the spectral power in the low theta bin (3 Hz) over the midline region is negatively correlated with operation span. A positive correlation with operation span is observed at central parietal and bilateral anterior temporal electrodes in the low alpha frequency bin (9 Hz).
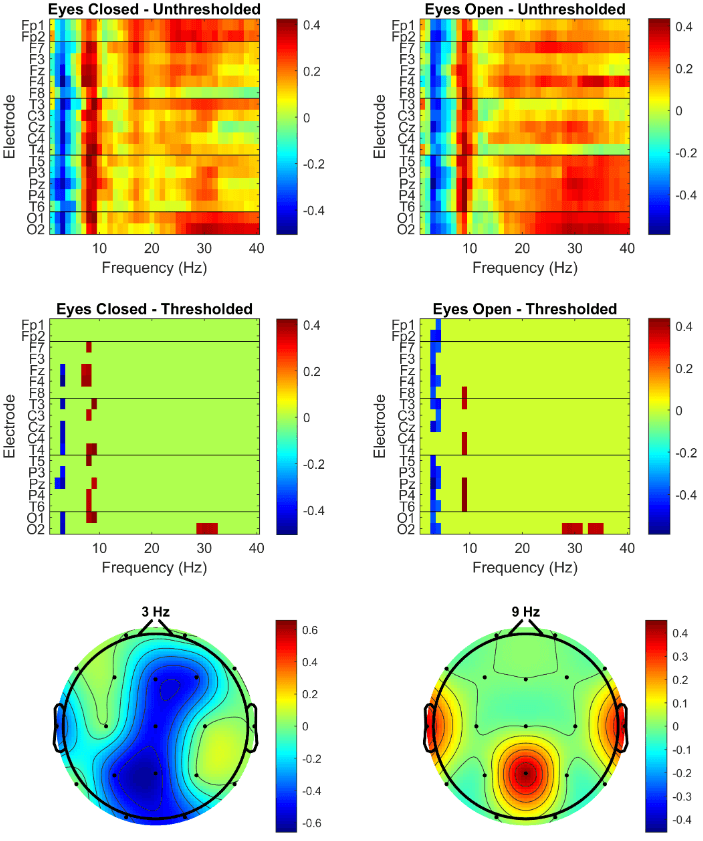
Figure 1: Correlational results. The top row shows the unthresholded correlations between the operation span scores and the relative spectral powers for each electrode-frequency pair in the eyes closed (left panel) and eyes open (right panel) conditions. The middle row presents only those correlations that survived the permutation test. The bottom row shows the topographic maps of the electrode-frequency pairs that remained above threshold in both the ocular conditions with both having an absolute correlation larger than 0.4.
Discussion
The aim of this study was to explore which brain wave oscillations in rest correlate with performance in the operation span task. The results overwhelmingly demonstrate that low theta and low alpha have negative and positive correlations with working memory capacity, respectively. Although this pattern is found across all electrodes in the uncorrected statistics, a more detailed pattern remained in the thresholded profile, which can be split into a midline-theta cluster and a parietotemporal-alpha cluster. The parietal location is consistent with previous neuroimaging findings. However, the novelty here is that the measurements were taken when the person was not engaged in any cognitive demanding task. An interpretation of why this region might be correlated with working memory capacity comes from its role in attention [23, 24].
For example, when participants pay more attention, the power in the alpha band increases [24]. Greater amount of attention paid to information is necessary for its entry into working memory. Whereas the increase in alpha is logically correlated with working memory capacity, the negative correlation with midline theta requires more consideration. Higher relative theta frequency necessarily means that the other (faster) frequencies are less prominent, including parietal alpha. The correlation between 3 Hz and 9 Hz at Pz is indeed negative (r = -0.48, p < 0.01). Given that parietal and frontal regions are connected via the default mode network, it suggests that greater relative alpha spectral power pushes the network into a higher level of attentional preparedness at the cost of lowering the relative theta power in the network and therefore lower ability of information integration [25].
During an actual working memory task, the balance will shift such that theta power increases to support the internal processing and alpha decreases to block any distracting information. The central parietal region therefore functions as a switch between outward and inward attention, using the alpha/theta balance as its currency. Individual differences in the alpha/theta balance are therefore associated with working memory capacity. To wit, given that correlation does not mean causation, the reverse interpretation can also be made, i.e., individual differences in working memory capacity could reflect differences in setpoints of the balance between outward attention at the expense of internal processing. As this situation is at rest, during task performance there is more cognitive room to increase internal processing and decrease external distraction and thus higher performance on the operation span task. This reconciles the contrasting correlational findings at rest and during task performance. Future studies could investigate whether individuals who learned to increase alpha relative to theta improve in operation span performance and in doing so their working memory capacity.
The current study used data from an existing study and therefore was not optimised for addressing more fine-grained questions. For example, this approach could be used with several measures of working memory capacity to extract a task-independent latent factor of capacity. A further extension would be to increase the electrode density or even replicate this approach using magnetoencephalography. The aim remains the same, which is to take a measure of working memory capacity and ask what brain pattern in rest is associated with it. The current findings suggest a much closer link between attentional preparedness and working memory than was previously thought and warrants further exploration.
Acknowledgements
The data were collected with funding from the Wellcome Trust (ref 204770/z/16/z).
Article Info
Article Type
Research ArticlePublication history
Received: Wed 08, Jul 2020Accepted: Sat 18, Jul 2020
Published: Tue 25, Aug 2020
Copyright
© 2023 Eddy J. Davelaar. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. Hosting by Science Repository.DOI: 10.31487/j.NNB.2020.03.05
Figures & Tables
References
- Miyake A, Shah P (1999) Models of working memory: Mechanisms of active maintenance and executive control. Cambridge Univ Press.
- Baddeley A (1986) Working Memory. New York: Oxford Univ Press.
- Barouillet P, Bernardin S, Portrat S, Vergauwe E, Camos V (2007) Time and cognitive load in working memory. J Exp Psychol Learn Mem Cogn 33: 570-585. [Crossref]
- Raaijmakers JG, Shiffrin RM (1981) Search of associative memory. Psychol Rev 88: 93-134.
- Altmann EK, Schunn CD (2012) Decay versus interference: a new look at an old interaction. Psychol Sci 23: 1435-1437.
- Cowan N (1995) Attention and Memory: An Integrated Framework. New York: Oxford Univ Press.
- Farrell S (2012) Temporal clustering and sequencing in short-term memory and episodic memory. Psychol Rev 119: 223-271. [Crossref]
- Henson RNA Burgess N, Frith CD (2000) Recoding, storage, rehearsal and grouping in verbal short-term memory: an fMRI study. Neuropsychologia 38: 426-440. [Crossref]
- Jonides J, Schmacker EH, Smith EE, Lauber EJ, Awh E et al. (1997) Verbal working memory load affects regional brain activation as measured by PET. J Cogn Neurosci 9: 462-475. [Crossref]
- Paulesu E, Frith CD, Frackowiak RSJ (1993) The neural correlates of the verbal component of working memory. Nature 362: 342-345. [Crossref]
- Shivde G, Thompson Schill SL (2004) Dissociating semantic and phonological maintenance using fMRI. Cogn Affect Behav Neurosci 4: 10-19. [Crossref]
- Luck SJ, Vogel EK (1997) The capacity of visual working memory for features and conjunctions. Nature 390: 279-281. [Crossref]
- Vogel EK, Machizawa MG (2004) Neural activity predicts individual differences in visual working memory capacity. Nature 428: 748-751. [Crossref]
- Luria R, Balaban H, Awh E, Vogel EK (2016) The contralateral delay activity as a neural measure of visual working memory. Neurosci Biobehav Rev 62: 100-108. [Crossref]
- Turner M, Engle RW (1989) Is working memory capacity task dependent? J Mem Lang 28: 127-154.
- Osaka N, Osaka M, Kondo H, Morishita M, Fukuyama H et al. (2004) The neural basis of executive function in working memory: an fMRI study based on individual differences. Neuroimage 21: 623-631. [Crossref]
- Scharinger C, Soutschek A, Schubert T, Gerjets P (2017) Comparison of the Working Memory Load in N-Back and Working Memory Span Tasks by Means of EEG Frequency Band Power and P300 Amplitude. Front Hum Neurosci 11: 6. [Crossref]
- Chuang K, Chen Y, Balachandran P, Liang W, Juan C (2019) Revealing the Electrophysiological Correlates of Working Memory-Load Effects in Symmetry Span Task With HHT Method. Front Psychol 10: 855. [Crossref]
- Gevins A, Smith ME, McEvoy L, Yu D (1997) High-resolution EEG mapping of cortical activation related to working memory: effects of task difficulty, type of processing, and practice. Cereb Cortex 7: 374-385. [Crossref]
- Jensen O, Gelfand J, Kounios J, Lisman JE (2002) Oscillations in the alpha band (9-12 Hz) increase with memory load during retention in a short-term memory task. Cereb Cortex 12: 877-882. [Crossref]
- Davelaar EJ, Jilek J (2020) Sensorimotor rhythm is associated with reinforcement learning and cognitive impulsivity: a neurofeedback study. Curr Neurobiol.
- Unsworth N, Heitz RP, Schrock JC, Engle RW (2005) An automated version of the operation span task. Behav Res Methods 37: 498-505. [Crossref]
- Benedek M, Schickel RJ, Jauk E, Fink A, Neubauer AC (2014) Alpha power increases in right parietal cortex reflects focused internal attention. Neuropsychologia 56: 393-400. [Crossref]
- Klimesch W (2012) α-band oscillations, attention, and controlled access to stored information. Trends Cogn Sci 16: 606-617. [Crossref]
- Alves PN, Foulon C, Karolis V, Bzdok D, Margulies DS et al. (2019) An improved neuroanatomical model of the default-mode network reconciles previous neuroimaging and neuropathological findings. Commun Biol 2: 370. [Crossref]