Depigmenting Effect of Azelaic Acid and Glycolic Acid in MNT-1 and B16-F10 Melanoma Cells
A B S T R A C T
Treatment for abnormal melanin accumulation is available with controversial results, thus aim of the present work was to determine azelaic acid (AZ) and glycolic acid (GLY) cytotoxicity and inhibiting melanogenesis efficacy. Safety concentration and efficacy in reducing the melanin content were access through MTT assay, Fontana-Masson staining, melanin content assay and tyrosinase (TYR) assay. Kojic acid (KO) was used as the positive control throughout the assays. AZ and GLY exhibited low cytotoxicity at 100 µg/mL and do not show suppression towards cell viability as cells were 100% viable. 100 µg/mL and 50 µg/mL AZ and GLY show comparable depigment effect to the positive control in Fontana-Masson staining and the depigmentation effect was dose dependent. 100 µg/mL AZ and 100 µg/mL GLY shows significant (P ≤ 0.05) reduction of melanin content in both melanoma cells compared to untreated group. TYR activity in both cells also shows significant lower (P ≤ 0.001) after the treatment with 100 µg/mL AZ and 100 µg/mL GLY compared to untreated group. AZ and GLY were suggested to suppress melanin formation by directly inhibiting TYR activity and act as TYR inhibitor. The depigmenting effect was further confirmed with down-regulating of TYR, TYRP1 and TYRP2. The possible depigmentation mechanisms of AZ and GLY could be due to inhibition of melanin synthesis and suppression of tyrosinase-related genes expression including which affecting TYR activity. The depigmenting potential of AZ and GLY can be evaluate further in-vivo model.
Keywords
Azelaic acid, glycolic acid, depigmentation, tyrosinase inhibitor, melanogenesis
Introduction
Melanin is the colour pigment which contributes to skin colour in addition to its protective properties against harmful ultraviolet radiation and oxidative stresses [1]. However, melanin accumulation can cause aesthetic problem such as melasma, freckles, senile lentigines and periorbital hyperpigmentation (POH) [1-4]. The production of melanin in melanocytes is predominantly determined by tyrosinase (TYR) regulation. The first step and the only rate-limiting step of melanin synthesis is the conversion of tyrosine to dihydroxyphenylalanine (DOPA) and dopaquinone by TYR [5]. This step is common to both forms of melanin red pheomelanin and the more abundant brown black eumelanin production. From there, synthesis of eumelanin requires further oxidation of dopa to dopachrome, followed by a series of oxidation reactions from dihydroxyindole and dihydroxyindole-2-carbolic acid [6]. While in the presence of cysteine or glutathione, dopauinone is converted to cysteinyldopa or glutothionyldopa. Subsequently, phenomelanin is formed [6].
There are several signaling pathways involved in melanin biosynthesis. Microphthalmia transcription factor (MITF) is a tissue restricted, basic helix-loop-helix leucine zipper, dimeric transcription factor which act as a master regulator of melanocytes development [7, 8]. MITF is known to regulate the transcription of TYR, tyrosinase related protein 1 (TYRP1), and tyrosinase related protein 2 (TYRP2) [9-12]. Binding of α-melanophore stimulating hormone (α-MSH) to melanocortin 1 receptor (MC1R) on melanocytes activates adenylate cyclase to produce cAMP as an intracellular second messenger via G-protein-coupled receptor type activation. cAMP binds and activates protein kinase A, which then activates the expression of MITF via phosphorylation of the cAMP response element-binding protein (CREB). Therefore, the use of small molecules that cease the signaling pathways involved in melanin biosynthesis and transformation might be useful in developing a new class of depigmenting agents for cosmetic purposes.
Kojic acid has been widely used as a positive control in the literature to compare the inhibitory strength of the targeted products in-vitro [13, 14]. Although kojic acid was shown to be a good TYR inhibitor, but it has some undesirable side effects that similar to hydroquinone such as erythema and scaling [15]. Thus, there is a need to look for an alternative of depigmentation agent. In industrial point of view, cosmetics can help to depigment skin colour, which is one of the important parts among the cosmetic fields. Particularly, development of whitening phytochemicals from natural sources has become new popular trends recently. The study of azelaic acid (AZ) on hyperpigmentation was coined when researchers notice the inhibiting effect of AZ on TYR [16]. Mechanism of action is through DNA synthesis and mitochondrial enzyme inhibition which results in inducing a direct cytotoxic effect towards melanocytes and cell proliferation [16-18]. Glycolic acid (GLY) is a versatile peeling agent used to treat defects of the epidermis and papillary dermis in the cosmetic and beauty industry [19].
Study done by Usuki et al. to identify the direct effect of GLY on melanogenesis using B16 and human melanoma cells found that glycolic acid can act as a competitive inhibitor to TYR; inhibits melanin formation without affecting the cell growth [20]. In most of the studies, combination with other active ingredient such as hydroquinone, AZ and KO give a more promising to reduce pigmentation and melasma [21, 22]. GLY plays an important role in shortening the cell cycle, facilitates depigmentation, interferes with pigment transfer, and enhances penetration of another depigmenting agent [23]. Although, the depigmenting ability of AZ and GLY were reported widely but the mechanism of both to cause the depigmentation was not studied comprehensively. In this present study, we demonstrated that the AZ and GLY are effective in depigmentation by downregulating the melanogenic regulatory genes and their corresponding transcription factors including MITF.
Materials and Methods
I Cell Culture
Two different melanoma cells were involved in the study, MNT-1 cells, a highly pigmented Homo sapiens melanoma cells and Mus musculus melanoma cells, B16-F10. MNT-1 cells, which acquired as gift from Dr. Michael S. Marks (University of Pennsylvania) were cultured in a mix medium of Dulbecco’s modified Eagle’s medium (DMEM) and Aim-V® medium with 20% (v/v) fetal bovine serum, 1% (v/v) non-essential amino acid, and 1% (v/v) sodium pyruvate. While B16-F10 cells, which purchased from the American Type Culture Collection (ATCC) were cultured in DMEM with 10% (v/v) fetal bovine serum, 1% (v/v) sodium pyruvate, and 1% (v/v) streptomycin and penicillin. Both of the cells were incubated at 37°C in humidified 5% CO2, 95% air.
II MTT Assay
Cell viability was measured by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assays following a 72-hour incubation according to the modified method of Carmichael et al. (1987). Briefly, 90 µl/well of cell suspension (5 × 103 cells/well) was added to the 96-well microculture plates. The cells were treated with 10 µl of AZ and GLY in seven ascending dose concentrations from 3.125, 6.25, 12.5, 25, 50, 100 and 200 µg/ml dissolved in dimethyl sulfoxide (DMSO). The cells were treated with DMSO and cell culture media, which served as the vehicle and negative control. After 24 hours of incubation, MTT solution (Sigma, St. Louis, MO, USA; 50 µl, 2 mg/ml) was added to each well 4h before termination. After 4 hours of incubation, the supernatant has removed and added with DMSO. The plate was analysed on ELISA plate reader (BioTek, United State) at 570nm.
III Fontana-Masson Stain
Based on the results that obtained from the MTT assay, at the end of cell culture with the treatment, the cells were fixed with 3.7% formaldehyde solution. Fontana-Masson staining of cells was conducted using a Fontana-Masson stain Kit (Polysciences, Inc., Warrington, UK, England) according to the manufacturer’s instructions. Briefly, the fixed cells were treated with working silver solution, 0.1% gold chloride solution, 5% sodium thiosulfate followed by nuclear-fast red solution. Fontana-Masson Stain Kit was used in the visualization of argentaffin granules and melanin, where cell nuclei, melanin and argentaffin would display red and black respectively, after staining. The staining cells will be photographed under a digital camera microscopy (Carl Zeiss, United States).
IV Melanin Content Quantification
AZ and GLY will only involve in the following experiment. Melanin content was measured according to the modified method of Uchida et al. In brief, 3 × 105 cells/well were treated for 24 hours with phytochemicals. Untreated cells served as controls. Cells were collected by trypsinizing them and centrifuged at 3,000g for 20 minutes. After precipitation, cells were resuspended in 200 μl of 2M NaOH incubated at 60°C for 20 minutes. Spectrophotometric analysis of melanin content was performed at 450nm absorbance (BioTek, United State). Synthetic melanin (Sigma, St. Louis, MO, USA) was used as a standard (Halaban et al. 1983, Naeyart et al. 1991).
V Tyrosinase Assay
The relative amounts of tyrosinase in treated cells and untreated cells were elucidated in this stage through tyrosinase expression profile engendered using Tyrosinase Colorimeteric Cell-Based ELISA Kit (CytoGlow, United State). In a 96-well platform, 1 × 104 cells/well were seeded overnight at 37ºC in CO2 incubator for treated with the phytochemicals the following day. At the end of the cell culture with the treatment of phytochemicals, the medium was removed and rinsed twice with 200 µL/well of 1 × TBS. 100 µL/well of 4% formaldehyde solution was added with incubation of 20 minutes in room temperature to fix the cells. Thereafter, the liquid content in each of the well was discarded, washed 3 times with 200 µL washed buffer for five minutes each with gentle shaking on orbital shaker. The plate was proceeded with tyrosinase assay according to the manufacturer’s instructions.
VI Melanogenesis Elucidation
1 × 107 of treated and untreated MNT-1 melanoma cells were harvested for extraction of RNA using RNA extraction kit. Reverse transcription polymerase chain reaction was carried out with High-Capacity RNA-to-cDNA kit (Applied Biosystems, USA). 2 µg of total RNA were used in the conversion of RNA-to-cDNA in a 20 µL reaction. The reverse transcription was performed using thermal cycler at the condition 37 ºC for 60 minutes and stopped by heating up at 95 ºC for 5 minutes. PCR Array was utilized to determine the level of mRNA expression that associated with melanogenesis. The genes which have been quantified were MITF, TYR, TYRP1 and TYPR2. While actin (ACTB) and glycerealdehyde-3-phosphate dehydrogenase (GADPH) acts as the endogeneous control. All of these genes were purchase from Applied Biosystem with gene code Hs01117294_m1 (MITF), Hs00267167_s1 (MC1R), Hs00165976_m1 (TYR), Hs00167051_m1 (TYRP1), Hs01098278_m1 (TYRP2), Hs99999903_m1 (ACTB) and Hs99999905_m1 (GADPH) respectively. The template cDNA was diluted to 4× from 100 ng/µl cDNA stock.
Quantitative polymerase chain reaction (qPCR) was performed using StepOnePlus Real-Time PCR System (Applied Biosystems, USA). qPCR was carried out in 20 µl reaction mixture consisting of 2 µl of 4× diluted template cDNA, 1 µl of Taqman® Gene Expression Assay and Taqman® Fast Advanced Master Mix (Applied Biosystems, USA). The sample was first denatured at 95 °C for 20 s, followed by 40 cycles of denaturing step at 95 °C for 1 s and annealing step at 60 °C for 20 s. The data was collected at the end of each annealing step.
Results
I MTT Assay
To identify the non-cytotoxic concentration of the phytochemicals, we treated MNT-1 and B16-F10 with phytochemicals at various concentrations; cell viability was determined using MTT assay. Figure 1A illustrates the cell viability profile of B16-F10 melanoma cells. 100% viability of B16-F10 were observed with AZ and GLY treated at doses ranging from 3.125 µg/mL to 100 µg/mL, Similar trend was observed as shown in (Figure 1B) when MNT-1 were treated AZ and GLY. MNT-1 were 100% viable when treated with AZ and GLY at doses ranging from 3.125 µg/mL to 100 µg/mL. Taken together, the maximal concentration to be used in vitro for further evaluation were 100 µg/mL of AZ and 100 µg/mL of GLY.

Figure 1: Dose-response curves of cell lines treated with four phytochemicals. A) B16-F10 melanoma cells were treated at seven concentration range of 2-fold dilution at 3.25-200 μg/mL with AZ and GLY for 72 hours. B) MNT-1 melanoma cells were treated at seven concentration range of 2-fold dilution at 3.25-200 μg/mL with AZ and GLY for 72 hours. The curve is expressed as mean ± SD.
II Evaluation of Depigmenting Activity
i Fontana Masson Stain
Both melanoma cells were treated with the maximal range of non-cytotoxic concentration of the phytochemicals and the melanin content of the respective treatments were observed by using Fontana Masson staining of the cells and qualitative monitored under inverted light microscope. After three passages the B16-F10 and MNT-1 turned black symbolize the production of melanin. KO at concentration 100 µg/mL was act as positive control [24]. MNT-1 which is highly pigmented appears to be darker compared to B16-F10. What stand out between all treatment groups and untreated in both melanoma cells were, treated groups exhibit nuclei in pink colour which is concur to KO at concentration 100 µg/mL. The degrees of pigmentation in melanoma cells were lighter and lower in amount as compared to untreated control. Of note, with successive increasing in non-cytotoxic concentration of phytochemicals, the degree of pigmentation decreases inverse proportionally showed that the depigmentation effect is dose dependent. Among the four phytochemicals, cells treated with 100 µg/mL and 50 µg/mL AZ and GLY show comparable depigment effect to the positive control in both melanoma cells. The results of the B16-F10 melanoma cells staining are shown in (Figures 2 & 3) shows the staining results of MNT-1 melanoma cells.

Figure 2: Fontana Masson staining on B16-F10 melanoma cells with respective phytochemicals. Melanin stained black and nuclei stained with pink. Staining images were taken with inverted light microscope under magnification of 100×. A) 100 µg/mL azelaic acid; B) 50 µg/mL µg/mL azelaic acid; C) 25 µg/mL azelaic acid; D) 100 µg/mL glycolic acid; E) 50 µg/mL glycolic acid; F) 25 µg/mL glycolic acid; G) Untreated; H) Positive control 100 µg/mL kojic acid.

Figure 3: Fontana Masson staining on MNT-1 melanoma cells with respective phytochemicals. Melanin stained black and nuclei stained with pink. Staining images were taken with inverted light microscope under magnification of 100×. A) 100 µg/mL azelaic acid B) 50 µg/mL µg/mL azelaic acid; C) 25 µg/mL azelaic acid; D) 100 µg/mL glycolic acid; E) 50 µg/mL glycolic acid; F) 25 µg/mL glycolic acid; G) Untreated; H) Positive control 100 µg/mL kojic acid.
ii Melanin Content Quantification
Three ascending concentrations 25 µg/mL, 50 µg/mL and 100 µg/mL of AZ and Gly were selected to further identify which grants the significant depigmenting effect in reducing melanin concentration in the melanoma cells. Treatment of 100 µg/mL of AZ, melanin content of B16-F10 (21.4 ± 1.95) and MNT-1 (197.2 ± 12.7) were significant lower (P ≤ 0.001) compared to untreated group in B16-F10 (36.9 ± 2.05) as well as in untreated group of MNT-1 (230.3 ± 15.7). Treatment with 50 µg/mL of AZ in MNT-1 (204.4 ± 16.7) was also significant lower (P ≤ 0.01) compared with untreated group as shown in (Figures 4A & 4B). Similar outcomes were observed in B16-F10 with 50 µg/mL AZ treatment group compared with untreated group. Figures 4C & 4D presents the summary outcomes of melanin content in both melanoma cells with GLY treatment. The melanin content in B16-F10 treated with 100 µg/mL GLY (31.9 ± 6.71) was significant lower (P ≤ 0.001) as compared to untreated group (40.1 ± 3.50). Besides that, 100 µg/mL of GLY (204.1 ± 20.1) treated on MNT-1 was also significant lower (P ≤ 0.001) compared with untreated (250.3 ± 3.83). 25 µg/mL and 50 µg/mL GLY showed similar results with the untreated group. Indeed, the effective concentration which lowers the melanin content in B16-F10 and MNT-1 were 100 µg/mL.
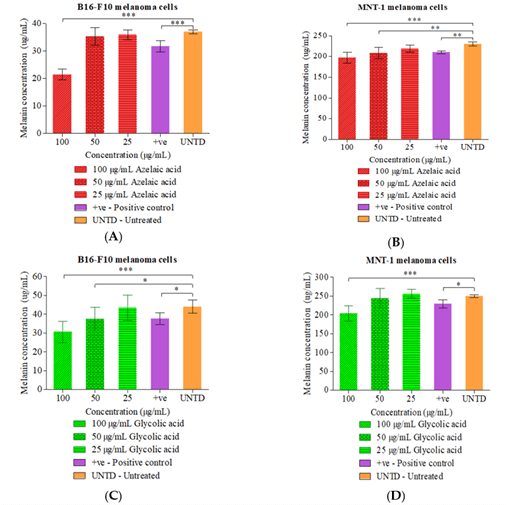
Figure 4: Melanin concentrations of cell lines after treated with respective phytochemicals at 2-fold dilution at 25-100 µg/mL. A) B16-F10 melanoma cells treated with azelaic acid; B) MNT-1 melanoma cells treated with azelaic acid; C) B16-F10 melanoma cells treated with glycolic acid; D) MNT-1 melanoma cells treated with glycolic acid.
iii Tyrosinase Assay
In order to assess the effect of AZ and GLY on TYR expression profile after the treatment, TYR assay was carried out. Three ascending concentrations 25 µg/mL, 50 µg/mL and 100 µg/mL of AZ and Gly were selected to further identify which grants the significant in reducing TYR activity in the melanoma cells. As showed in (Figure 5), TYR reduction was observed after treatment with AZ and GLY. Relative tyrosinase activity of the AZ treated group in B16-F10 (83.8 ± 4.71) and MNT-1 (85.6 ± 10.8) were significant lower (P ≤ 0.001) than untreated group in B16-F10 (100 ± 0), as well as in untreated group of MNT-1 melanoma cell (100 ± 0). On the other hand, the relative activity of TYR treated with GLY in B16-F10 (86.3 ± 8.45) and MNT-1 (83.6 ± 9.58) also shows significant lower compared to B16-F10 untreated group (100 ± 0) and MNT-1 untreated group (100 ± 0). Similar outcomes were observed among the treatment group with 50 µg/mL and 25 µg/mL of AZ and GLY and untreated group for both melanoma cells. 100 µg/mL of AZ and GLY were effective in lowering TYR activity.

Figure 5: Tyrosinase activity of cell lines after treated with respective phytochemicals at 2-fold dilution at 25-100 µg/mL. A) B16-F10 melanoma cells treated with azelaic acid; B) MNT-1 melanoma cells treated with azelaic acid; C) B16-F10 melanoma cells treated with glycolic acid; D) MNT-1 melanoma cells treated with glycolic acid.
III Melanogenesis Elucidation
The relative fold changes of expression or repression of each gene after 6- and 12-hour treatment compared to respective untreated group were calculated using the comparative CT method. Fold changes for MITF, MC1R, TYR, TYRP1 and TYRP2 are shown in (Table 1). The fold expression calculated using formula -ΔΔCT of each melanogenesis related genes after treatment with AZ and GLY are illustrated in (Figure 6). We speculate that AZ and GLY have inhibitory effect on transcriptional level as MITF, MC1R, TYR, TYRP1 and TYRP2 shows down-regulation compared to the untreated. At 6-hour treatment with 100 µg/mL of AZ, TYRP2 (69%) shows the highest down-regulation among all of the genes followed by MC1R (47%), TYR (39%), TYRP1 (22%) and lastly MITF (17%) compared to endogenous control. For 6-hour treatment with 100 µg/mL of GLY, TYRP2 (69%) once again shows the highest down-regulation among all of the genes follow by TYR (61%), TYRP1 (41%), MITF (40%) and lastly MC1R (39%) compared to endogenous control.
Table 1: The expression of selected genes in AZ and GLY treated for 6- and 12-hour treatment calculation using-ΔΔCT.
|
Genes |
Groups |
Average CT ± SD |
Fold changes (2^- ΔΔCT) |
Fold differences (-ΔΔCT) |
|
MITF |
6 hour AZ treated |
18.3 ± 0.318 |
0.831 |
-0.265 |
|
12 hour AZ treated |
17.5 ± 0.216 |
0.527 |
-0.921 |
|
|
6 hour GLY treated |
18.8 ± 0.394 |
0.577 |
-0.792 |
|
|
12 hour GLY treated |
17.3 ± 0.241 |
0.607 |
-0.719 |
|
|
Untreated (6 hour) |
18.6 ± 0.273 |
1 |
0 |
|
|
Untreated (12 hour) |
17.5 ± 0.0847 |
1 |
0 |
|
|
MC1R |
6 hour AZ treated |
22.0 ± 0.379 |
0.880 |
-0.182 |
|
12 hour AZ treated |
22.6 ± 0.374 |
0.430 1 |
-1.21 |
|
|
6 hour GLY treated |
22.5 ± 0.216 |
0.615 |
-0.700 |
|
|
12 hour GLY treated |
22.5 ± 0.200 |
0.434 1 |
-1.20 |
|
|
Untreated (6 hour) |
22.4 ± 0.363 |
1 |
0 |
|
|
Untreated (12 hour) |
22.3 ± 0.594 |
1 |
0 |
|
|
TYR |
6 hour AZ treated |
21.8 ± 0.183 |
0.533 |
-0.906 |
|
12 hour AZ treated |
22.2 ± 0.234 |
0.337 1 |
-1.56 |
|
|
6 hour GLY treated |
22.3 ± 0.184 |
0.391 1 |
-1.35 |
|
|
12 hour GLY treated |
22.0 ± 0.215 |
0.389 1 |
-0.389 |
|
|
Untreated (6 hour) |
21.5 ± 0.218 |
1 |
0 |
|
|
Untreated (12 hour) |
21.6 ± 0.214 |
1 |
0 |
|
|
TYRP1 |
6 hour AZ treated |
19.5 ± 0.313 |
0.612 |
-0.706 |
|
12 hour AZ treated |
20.2 ± 0.429 |
0.335 1 |
-1.57 |
|
|
6 hour GLY treated |
19.6 ± 0.235 |
0.590 |
-0.760 |
|
|
12 hour GLY treated |
19.7 ± 0320 |
0.452 1 |
-1.14 |
|
|
Untreated (6 hour) |
19.4 ± 0.104 |
1 |
0 |
|
|
Untreated (12 hour) |
19.5 ± 0.260 |
1 |
0 |
|
|
TYRP2 |
6 hour AZ treated |
22.7 ± 0.139 |
0.311 1 |
-1.68 |
|
12 hour AZ treated |
21.9 ± 0.109 |
0.385 1 |
-1.37 |
|
|
6 hour GLY treated |
22.7 ± 0.222 |
0.311 1 |
-1.68 |
|
|
12 hour GLY treated |
21.3 ±0.248 |
0.569 |
-0.811 |
|
|
Untreated (6 hour) |
21.6 ± 0.340 |
1 |
0 |
|
|
Untreated (12 hour) |
21.5 ± 0.0865 |
1 |
0 |

Figure 6: Differential expressions of selected genes of azelaic acid and glycolic acid treated in compared with untreated control. MNT-1 melanoma cells were treated at 100 μg/mL of AZ and 100 μg/mL of GLY for 6 hours and 12 hours respectively. Treated group cellular expression of MITF, MC1R, TYR, TYRP1 and TYRP2 were compared to the respective hour untreated group. ACTB and GADPH act as the endogenous control and normalized with MITF, MC1R, TYR, TYRP1 and TYRP2.
When prolong to 12-hour incubation, both phytochemical exhibit a better repression on the genes. Melanoma cells treated with 100 µg/mL of AZ for 6 hour shows 90% of down regulation on TYRP2 followed by TYR (67%), TYRP1 (67%), MC1R (57%) and MITF (48%) compared to endogenous control. While 100 µg/mL of GLY also shows a better repression of genes where TYRP2 were downregulated by 76% compared to endogenous control follow by TYR (62%), MC1R (57%), TYRP1 (55%) and MITF (43%). Significant down-regulation was confirmed with the fold change between 0 to 0.49, while fold change between 0.5 to 1 means no difference in treated and untreated groups. Significant down-regulation was only observed at TYRP2 with AZ at 6-hour treatment, while MC1R, TYR, TYRP1 and TYRP2 at 12-hour treatment compared to untreated group. Treatment with GLY shows significant down-regulation in TYR, TYRP1 and TYRP2 at 6- and 12-hour incubation compared to untreated group.
Discussion
Among the possible melanin reducing compounds, TYR inhibitor is the most effective way to treat pigmentation and are used as skin-whitening agents in the cosmetic industry [25]. AZ was first discovered as the competitive inhibitors in mushroom TYR [26, 27]. Several studies reported that AZ can block TYR activity as competitive inhibitor and DNA synthesis in melanoma cell lines [17, 28]. We speculate AZ can be a TYR inhibitor as demonstrated earlier, 100 µg/mL AZ were able to shows comparable depigment effect to 100 µg/mL of KO (positive control), significantly decrease in melanin concentration (P ≤ 0.001) and TYR activity (P ≤ 0.001) were observed in both melanoma cells. Our results support evidence from previous observations that showed TYR activity was decreased with the treatment of AZ [17, 28]. Our results are also consistent with Yu and Kim study which demonstrated that AZ alone has the inhibitory effects in B16-F10 without significant cytotoxicity. Yu and Kim also further reported that the combination of AZ and taurine showed better depigmenting effect compared to AZ alone [29]. Thus, we suggest that the reduction of cellular melanin content in both melanoma cells were due to competitive inhibition on TYR where AZ prevent the binding of TYR to tyrosine, DOPA and dihydroxyindole.
GLY is well known as versatile peeling agent which able to accelerate the turnover of cells result in skin lightening. However, not much study was carried out to evaluate the inhibition of GLY on melanin production. Lim and Tham reported that GLY can be used to treat hyperpigmentation [22]. Our results speculate GLY can be TYR inhibitor which is in line with previous studies [20, 30, 31]. Determination of melanin synthesis in MNT-1 and B16-F10 showed that melanin content was significantly decreased (P ≤ 0.001) with GLY treatment. This inhibition effect of GLY could be attributed to the inhibition of TYR activity which was significantly decreased (P ≤ 0.001) with GLY treatment. Usuki and colleague reported that GLY inhibit TYR enzyme activity directly on human melanoma cells and the effect is independent of their acidic nature [20]. In our study, it is likely that GLY could be suppressing melanin formation by directly inhibiting TYR activity too as our results are in line with Usuki and colleague findings [20].
TYR plays a key role in production of melanin, because it catalyzes the rate-limiting steps involved in melanogenesis: the hydroxylation of tyrosine to 3,4-dihydroxyphenylalanine (DOPA), oxidation of DOPA, and conversion of 5,6-dihydroxyindole (DHI) to indole-5, 6-quinone [32]. We contemplated AZ and GLY as a TYR inhibitor where after the treatment, the degrees of pigmentation become lighter in Fontana Masson staining, melanin concentration and TYR activity were also significantly decreases (P ≤ 0.001) compared to untreated cell lines. AZ and GLY blocked the binding site of tyrosine which led to decreases in hydroxylation of tyrosine, oxidation of DOPA and conversion of 5,6-dihydroxyindole (DHI) to indole-5, 6-quinone thus prevent the production of eumelanin.
MITF is the master regulator of melanogenesis, which up-regulate the TYR, TYRP1 and TYRP2 via binding to the M-box motif in their promoter regions [27, 28]. The over expression of TYR, TYRP1 and TYRP2 will led to hyperpigmentation. Thus, the inhibition of melanogenesis related gene and protein expression determined the efficacy of depigmenting agents. Down regulation of these genes resulted in inhibition of melanogenesis as shown by decreased in TYR activity and decreased melanin content in skin melanocytes. Our results demonstrated that AZ treatment down-regulated TYR, TYRP1 and TYRP2 mRNA expression. These results suggested that AZ was able to decrease melanin production by inhibiting the expression of tyrosinase-related genes which also affecting the TYR activity as shown results of TYR assay. Similar findings were observed in a study which used anemonin, a natural compound found in plants. Anemonin was found to inhibit melanin synthesis by inhibiting the transcription of TYR, TYRP1 and TYRP2 [33].
So far, no study was done on how GLY affects the melanogenic gene expression. In this study, we found that GLY were involved in modulating the expression of TYR and tyrosinase-related-protein genes indicating their potential as whitening compound. These results suggest that GLY were able to decrease melanin production by inhibiting the expression of tyrosinase-related genes which also affecting the TYR activity and attribute to decrease in melanin production. Although the amount of melanin produced by melanocytes is strictly dependent on the level of TYR activity, the level of TYR activity in the cell is not dependent on the amount of enzyme produced. Melanocytes with high TYR activity do not necessarily attributed high transcription of TYR mRNA; in fact, it might be due to the regulated at a post-translational level [34]. The hormonal stimulation by α-MSH mediated through MC1R results in increased intracellular cAMP levels and leads to melanin synthesis [35]. Melanocortines (α-MSH and ACTH) can up-regulate expression of the MC1R, thus acting in a positive feedback loop [36]. Down-regulation of MC1R with the treatment with AZ and GLY decrease the production of MC1R, limit the binding of melanocortines thus prevent the differentiation of melanocytes.
Conclusion
The possible depigmentation mechanisms of AZ and GLY could be due to inhibition of melanin synthesis and suppression of tyrosinase-related genes expression including which affecting TYR activity. The depigmenting potential of AZ and GLY can be evaluate further in-vivo model.
Author Contributions
Conceptualization: Shiau-Chuen Cheah; Data curation: Kam Weng Chia; Formal analysis: Kam Weng Chia; Funding acquisition: Shiau-Chuen Cheah; Investigation: Kam Weng Chia; Methodology: Shiau-Chuen Cheah; Project administration: Shiau-Chuen Cheah; Resources: Siew Keah Lee and Shiau-Chuen Cheah; Supervision: Shiau-Chuen Cheah; Visualization: Kam Weng Chia; Writing-original draft: Kam Weng Chia; Writing-review and editing: Kam Weng Chia, Chung Keat Tan and Shiau-Chuen Cheah.
Funding
This research was funded UCSI University Research Grant Scheme [Proj-In-FMHS-021].
Acknowledgements
Many thanks to Ms Teoh Siew Hoon for her administrative support.
Conflicts of Interest
None.
Article Info
Article Type
Research ArticlePublication history
Received: Tue 01, Dec 2020Accepted: Fri 11, Dec 2020
Published: Fri 25, Dec 2020
Copyright
© 2023 Shiau-Chuen Cheah. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. Hosting by Science Repository.DOI: 10.31487/j.CEI.2020.03.05
Figures & Tables
Table 1: The expression of selected genes in AZ and GLY treated for 6- and 12-hour treatment calculation using-ΔΔCT.
|
Genes |
Groups |
Average CT ± SD |
Fold changes (2^- ΔΔCT) |
Fold differences (-ΔΔCT) |
|
MITF |
6 hour AZ treated |
18.3 ± 0.318 |
0.831 |
-0.265 |
|
12 hour AZ treated |
17.5 ± 0.216 |
0.527 |
-0.921 |
|
|
6 hour GLY treated |
18.8 ± 0.394 |
0.577 |
-0.792 |
|
|
12 hour GLY treated |
17.3 ± 0.241 |
0.607 |
-0.719 |
|
|
Untreated (6 hour) |
18.6 ± 0.273 |
1 |
0 |
|
|
Untreated (12 hour) |
17.5 ± 0.0847 |
1 |
0 |
|
|
MC1R |
6 hour AZ treated |
22.0 ± 0.379 |
0.880 |
-0.182 |
|
12 hour AZ treated |
22.6 ± 0.374 |
0.430 1 |
-1.21 |
|
|
6 hour GLY treated |
22.5 ± 0.216 |
0.615 |
-0.700 |
|
|
12 hour GLY treated |
22.5 ± 0.200 |
0.434 1 |
-1.20 |
|
|
Untreated (6 hour) |
22.4 ± 0.363 |
1 |
0 |
|
|
Untreated (12 hour) |
22.3 ± 0.594 |
1 |
0 |
|
|
TYR |
6 hour AZ treated |
21.8 ± 0.183 |
0.533 |
-0.906 |
|
12 hour AZ treated |
22.2 ± 0.234 |
0.337 1 |
-1.56 |
|
|
6 hour GLY treated |
22.3 ± 0.184 |
0.391 1 |
-1.35 |
|
|
12 hour GLY treated |
22.0 ± 0.215 |
0.389 1 |
-0.389 |
|
|
Untreated (6 hour) |
21.5 ± 0.218 |
1 |
0 |
|
|
Untreated (12 hour) |
21.6 ± 0.214 |
1 |
0 |
|
|
TYRP1 |
6 hour AZ treated |
19.5 ± 0.313 |
0.612 |
-0.706 |
|
12 hour AZ treated |
20.2 ± 0.429 |
0.335 1 |
-1.57 |
|
|
6 hour GLY treated |
19.6 ± 0.235 |
0.590 |
-0.760 |
|
|
12 hour GLY treated |
19.7 ± 0320 |
0.452 1 |
-1.14 |
|
|
Untreated (6 hour) |
19.4 ± 0.104 |
1 |
0 |
|
|
Untreated (12 hour) |
19.5 ± 0.260 |
1 |
0 |
|
|
TYRP2 |
6 hour AZ treated |
22.7 ± 0.139 |
0.311 1 |
-1.68 |
|
12 hour AZ treated |
21.9 ± 0.109 |
0.385 1 |
-1.37 |
|
|
6 hour GLY treated |
22.7 ± 0.222 |
0.311 1 |
-1.68 |
|
|
12 hour GLY treated |
21.3 ±0.248 |
0.569 |
-0.811 |
|
|
Untreated (6 hour) |
21.6 ± 0.340 |
1 |
0 |
|
|
Untreated (12 hour) |
21.5 ± 0.0865 |
1 |
0 |
References
- Goh MJ, Lee HK, Cheng L, Kong DY, Yeon JH et al. (2013) Depigmentation Effect of Kadsuralignan F on Melan-A Murine Melanosytes and Human Skin Equivalents. Int J Mol Sci 14: 1655-1666. [Crossref]
- Goding CR (2007) Melanocytes: the new Black. Int J Biochem Cell Biol 39: 275-279. [Crossref]
- Graziosi AC, Quaresma MR, Michalany NS, Ferreira LM (2013) Cutaneous Idiopathic Hyperchromia of the Orbital Region (CIHOR): a histopathological study. Aesthetic Plast Surg 37: 434-438. [Crossref]
- Slominski A, Tobin DJ, Shibahara S, Wortsman J (2004) Melanin Pigmentation in Mammalian Skin and Its Hormonal Regulation. Physiol Rev 84: 1155-1228. [Crossref]
- Chen XX, Shi Y, Chai WM, Feng HL, Zhuang JX et al. (2014) Condensed tannins from Ficus virens as tyrosinase inhibitors: structure, inhibitory activity and molecular mechanism. PloS One 9: e91809. [Crossref]
- Chang T (2012) Natural Melanogeensis Inhibitors Acting Through the Down-Regulation of Tyrosinase Activity. Materials 5: 1661-1685. [Crossref]
- Hodgkinson CA, Moore KJ, Nakayama A, Steingrímsson E, Copeland NG et al. (1993) Mutations at the mouse microphthalmia locus are associated with defects in a gene encoding a novel basic-helix-loop-helix-zipper protein. Cell 74: 395-404. [Crossref]
- Levy C, Khaled M, Fisher DE (2006) MITF: master regulator of melanocyte development and melanoma oncogene. Trends Mol Med 12: 406-414. [Crossref]
- Fang D, Setaluri V (1999) Role of microphthalmia transcription factor in regulation of melanocyte differentiation marker TRP-1. Biochem Biophys Res Commun 256: 657-663. [Crossref]
- Fang D, Kute T, Setaluri V (2001) Regulation of Tyrosinase‐related Protein‐2 (TYRP2) in Human Melanocytes: Relationship to Growth and Morphology. Pigment Cell Res 14: 132-139. [Crossref]
- Fang D, Tsuji Y, Setaluri V (2002) Selective down‐regulation of tyrosinase family gene TYRP1 by inhibition of the activity of melanocyte transcription factor, MITF. Nucleic Acids Res 30: 3096-3106. [Crossref]
- Hershey CL, Fisher DE (2005) Genomic analysis of the Microphthalmia locus and identification of the MITF-J/Mitf-J isoform. Gene 347: 73-82. [Crossref]
- Sato K, Morita M, Ichikawa C, Takahashi H, Toriyama M (2008) Depigmenting mechanisms of all-trans retinoic acid and retinol on B16 melanoma cells. Biosci Biotechnol Biochem 72: 2589-2597. [Crossref]
- Yao C, Oh JH, Oh IG, Park CH, Chung JH (2013) [6]-Shogaol inhibits melanogenesis in B16 mouse melanoma cells through activation of the ERK pathway. Acta Pharmacol Sin 34: 289-294. [Crossref]
- Nakagawa M, Kawai K, Kawai K (1995) Contact allergy to kojic acid in skin care products. Contact Dermatitis 32: 9-13. [Crossref]
- Halder RM, Richards GM (2004) Topical agents used in the management of hyperpigmentation. Skin Therapy Lett 9: 1-3. [Crossref]
- Detmar M, Mayer da Silva A, Stadler R, Orfanos CE (1989) Effects of azelaic acid on proliferation and ultrastructure of mouse keratinocytes in vitro. J Invest Dermatol 93: 70-74. [Crossref]
- Leibl H, Stingl G, Pehamberger H, Korschan H, Konrad K et al. (1985) Inhibition of DNA synthesis of melanoma cells by azelaic acid. J Invest Dermatol 85: 417-422. [Crossref]
- Murad H, Shamban AT, Premo PS (1995) The use of glycolic acid as a peeling agent. Dermatol Clin 13: 285-307. [Crossref]
- Usuki A, Ohashi A, Sato H, Ochiai Y, Ichihashi M et al. (2003) The inhibitory effect of glycolic acid and lactic acid on melanin synthesis in melanoma cells. Exp Dermatol 12: 43-50. [Crossref]
- Lim JT (1999) Treatment of melasma using kojic acid in a gel containing hydroquinone and glycolic acid. Dermatol Surg 25: 282-284. [Crossref]
- Lim JT, Tham SN (1997) Glycolic acid peels in the treatment of melasma among Asian women. Dermatol Surg 23: 177-179. [Crossref]
- Solano F, Briganti S, Picardo M, Ghanem G (2006) Hypopigmenting agents: an updated review on biological, chemical and clinical aspects. Pigment Cell Res 19: 550-571. [Crossref]
- Jamshidzadeh A, Shokri Y, Ahmadi N, Mohamadi N, Sharififar F (2017) Quercus infectoria and Terminalia chebula decrease melanin content and tyrosinase activity in B16/F10 cell lines. J Pharm Pharmacogn Res 5: 270-277.
- Liu SH, Pan IH, Chu IM (2007) Inhibitory effect of p-hydroxybenzyl alcohol on tyrosinase activity and melanogenesis. Biol Pharm Bull 30: 1135-1139. [Crossref]
- Nazzaro Porro M, Passi S (1978) Identification of tyrosinase inhibitors in cultures of Pityrosporum. J Invest Dermatol 71: 205-208. [Crossref]
- Niu C, Aisa HA (2017) Upregulation of Melanogenesis and Tyrosinase Activity: Potential Agents for Vitiligo. Molecules 22: 1303. [Crossref]
- Lemic Stojcevic L, Nias AH, Breathnach AS (1995) Effect of azelaic acid on melanoma cells in culture. Exp Dermatol 4: 79-81. [Crossref]
- Yu JS, Kim AK (2010) Effect of combination of taurine and azelaic acid on antimelanogenesis in murine melanoma cells. J Biomed Sci 17: S45. [Crossref]
- Badreshia Bansal S, Draelos ZD (2007) Insight into skin lightening cosmeceuticals for women of color. J Drugs Dermatol 6: 32-39. [Crossref]
- Bowe WP, Shalita AR (2008) Effective over-the-counter acne treatments. Frontline Medical Communications. Sem Cutan Med Surg 27: 170-176.
- Ito S, Wakamatsu K (2008) Chemistry of mixed melanogenesis-pivotal roles of dopaquinone. Photochem Photobiol 84: 582-592. [Crossref]
- Burchill SA, Virden R, Thody AJ (1989) Regulation of tyrosinase synthesis and its processing in the hair follicular melanocytes of the mouse during eumelanogenesis and phaeomelanogenesis. J Invest Dermatol 93: 236-240. [Crossref]
- Iozumi K, Hoganson GE, Pennella R, Everett MA, Fuller BB et al. (1993) Role of tyrosinase as the determinant of pigmentation in cultured human melanocytes. J Invest Dermatol 100: 806-811. [Crossref]
- Kondo T, Hearing VJ (2011) Update on the regulation of mammalian melanocyte function and skin pigmentation. Expert Rev Dermatol 6: 97-108. [Crossref]
- Horrell EMW, D’Orazio JA (2016) Melanocortin 1 Receptor (MC1R) as a Global Regulator of Cutaneous UV Responses: Molecular Interactions and Opportunities for Melanoma Prevention. Springer International Publishing. Skin Stress Response Pathways 155-174.
- Findlay GH, Morrison JG, Simson IW (1975) Exogenous ochronosis and pigmented colloid milium from hydroquinone bleaching creams. Br J Dermatol 93: 613-622. [Crossref]
- Koo JH, Kim HT, Yoon HY, Kwon KB, Choi IW et al. (2008) Effect of Xanthohumol on Melanogenesis in B16 Melanoma Cells. Exp Mol Med 40: 313-319. [Crossref]
- Pathak MA, Fitzpatrick TB, Kraus EW (1986) Usefulness of retinoic acid in the treatment of melasma. J Am Acad Dermatol 15: 894-899. [Crossref]